

janis
-
Постов
11 257 -
Зарегистрирован
-
Победитель дней
4
Сообщения, опубликованные janis
-
-
Pam Chin Остров Мбита, Замбия, озеро Танганьика
Камни здесь квадратные и покрыты улитками(Lavigeria и Parameliana sp.). Там, где я плавала, много прямого солнечного света и много корма-водорослей, и улитки это поняли.


Benoit Jonas : Есть довольно много видов, которые питаются улитками в озере, самые известные Neolamprologus tretocephalus, Chalinochromis, и N. mondabu тоже ...
У N. tretocephalus, зубы , чтобы размалывать панцири ,у Chalinochromis и N. mondabu, есть мощные клыки, чтобы извлечь тело из раковины. -
Новый вынашивающий потомство во рту вид Gymnogeophagus описан в притоке реки Уругвай.(Cichliformes: Cichlidae)
AAndréia Turcati, Wilson Sebastián ,Serra-Alanis Luiz, R. Malabarba.
Новый вид отличается от большинства видов рода наличием гипертрофированных губ, а от G. labiatus и G. pseudolabiatus - по цвету. Обсуждается наличие последовательных аллопатрических видов клады Gymnogeophagus gymnogenys, населяющей притоки реки Уругвай.

Голова Gymnogeophagus peliochelynion (первая колонка), G. pseudolabiatus (вторая колонка) и G. mekinos (третья колонка), показывающая полностью черный горб у самцов у G. peliochelynion (против желтого с черным краем), а верхняя губа не сложена дорсально по переднему краю морды (по сравнению с верхней губой, сложенной дорсально у G. pseudolabiatus и не развитой у G. mekinos). Gymnogeophagus peliochelynion сверху донизу, паратип, ZVC-P 13210, паратип, 76,3 мм SL; ZVC-P 7016, 89,9 мм SL; ZVC-P 13057, 90,2 мм SL. G. pseudolabiatus сверху вниз, паратип, UFRGS 7754, 102,0 мм SL; MHNM 4010, 88,8 мм SL; MHNM 4010, 95,3 мм SL. G. mekinos сверху вниз, MHNM 3511, 105,1 мм SL; MHNM 3511, 97,2 мм SL; MHNM 4009, 121,3 мм SL.
Окраска после фиксации в формалине. Зрелые самцы: основной цвет тела темно-коричневый над продольным рядом чешуек боковой линии и светло-желтовато-коричневый снизу. Ряд двойных вертикальных полосок отчетливо различим вдоль боковой поверхности , распределенных между основанием грудного плавника и концом хвостового стебля, спереди от основания хвостового плавника. Количество вертикальных двойных темных полос 5-6. Среднее пятно без определенных границ. Горб, полностью черный у самцов, переходит в темную полосу перед спинным плавником и едва заметна. Голова темно-коричневая с некоторыми черными пятнами на щеке; темная полоса, покрывающая щеку, едва различима. Перешеки и жаберно-пищеводные оболочки темно-коричневые. Две проксимальных трети лучей анального плавника покрыты маленькими круглыми точками. Хвостовой плавник покрыт белыми точками, за исключением верхней и нижней границ.
Цвет у сохранившихся самок и молодых особей не отличается от описанного для самцов. Основные отличия: перешеек и жаберная мембрана желтые; четко выраженная темная полоса, покрывающая щеку под глазом; отчетливая темная полоса на спинном плавнике; и больший размер белых пятен на спинных, анальных и хвостовых плавниках.

Gymnogeophagus peliochelynion : верх, голотип, самец, ZVC-P 12493, 101,9 SL; низ, паратип,самка, UFRGS 8076, 77,2 SL. Оба из Арройо-де-лас-Тунас на дороге 31, приток Рио-Арапей-Гранде, Сальто, Уругвай. Фотографии, сделанные сразу после сбора и фиксации в формалине.

Gelnogeophagus peliochelynion : выше, паратип, самец, ZVC-P 13210, паратип, 76,3 мм SL, Rio Rare, Colonia Lavalleja, Paso Elías, Salto,
Uruguay; ниже, самка, ZVC-P 13057, 65,3 мм SL, Arroyo Sopas, Paso del Cementerio, Salto, Uruguay. Фотографии живых образцов.
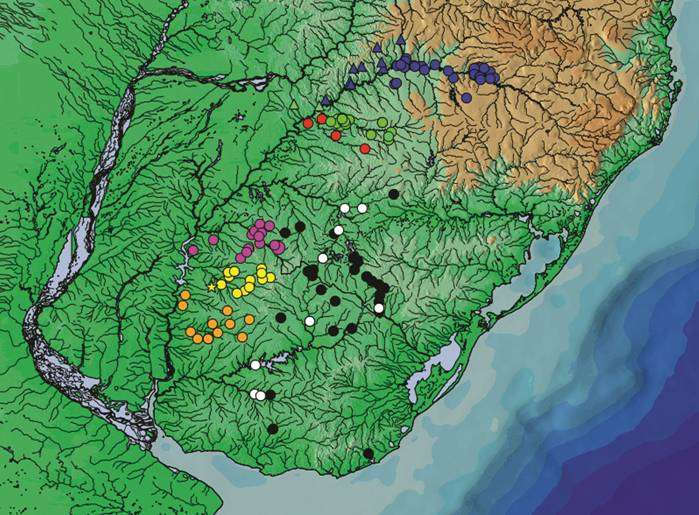
Географическое распространение. Этот вид известен по системе реки Арапей-Гранде, притока нижнего бассейна реки Уругвай, Уругвай
Виды клады Gymnogeophagus gymnogenys имеют аллопатрическое распределение вдоль нескольких притоков реки Уругвай . Gymnogeophagus lipokarenosэндемичен для верхней части реки Уругвай, вверх по течению до Сан-Хавьер (Аргентина) / Порто-Ксавьер (Бразилия). Gymnogeophagus constellatus и G . missioneiro обитают в притоках непосредственно ниже по течению от Сан-Хавьера и Порто-Ксавьера, первый встречается в основном в речных системах Рио-Июй (Бразилия), Арройо-Итакаруаре и Арройо-Чимирай-Мини (Мисьонес), а второй - в системе Рио-Пиратини (Бразилия), все из которых осушаются. базальтовая горная порода формации Серра Герал. Следующим основным притоком реки Уругвай в направлении вниз по течению является рио Ибикуи с мелкозернистым осадочным субстратом, соответствующим уникальному притоку среднего реки Уругвая, где были найдены G. tiraparae и G. mekinos . Эти два вида также присутствуют в нижних притоках бассейнов рек Уругвая и Ла-Платы в Уругвае и Бразилии. В следующем ниже по течению рио Quaraí, мы находим G. pseudolabiatus . Следующий приток ниже по течению, Рио Арапей, занят новым видом, описанным здесь, G. peliochelynion . Эта последовательная замена аллопатрических видов клады Gymnogeophagus gymnogenys вдоль притоков Рио-Уругвая предполагает, что основной канал действует как физический или экологический барьер для рассеивания этих видов, все из которых приспособлены к небольшим притокам. Аналогичные закономерности наблюдались в притоках реки Амазонки для других групп рыб и для прибрежных рек Атлантического океана, соединенных через пресноводные озера.
Популяции из Рио-Квейгай и Рио-Дейман, расположенные вниз по течению от Рио-Арапей, предварительно определены здесь как G. sp.peliochelynion . Хотя они не имеют гипертрофированных губ, они имеют окраску, подобную G. peliochelynion . Необходимы дальнейшие исследования, чтобы определить, составляют ли эти популяции отдельный вид.
Адаптированный перевод Aston S Martin
http://www.scielo.br/scielo.php?script=sci_arttext&pid=S1679-62252018000400218&lng=en&tlng=en
-
-
4 часа назад, sashh сказал:
Тут получается почти в 5 раз запас ))
вспомните меня когда у вас повится баночка на 600-700л...а она появится ))))))))
-
Lake Viktoria
Photo Lawrence Kent















-
35 минут назад, sashh сказал:
eheim classic 2260 для такого объема. чисто моё имхо-я бы взял с запасом, так сказать
-
Самец или самка? Как различить пол цихлид.
Гэри Краточвил.
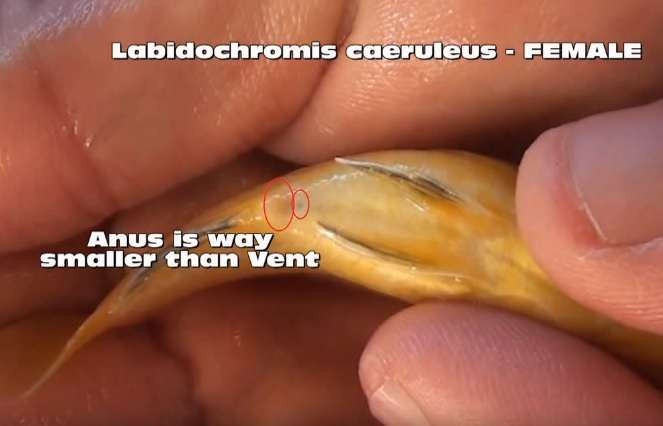
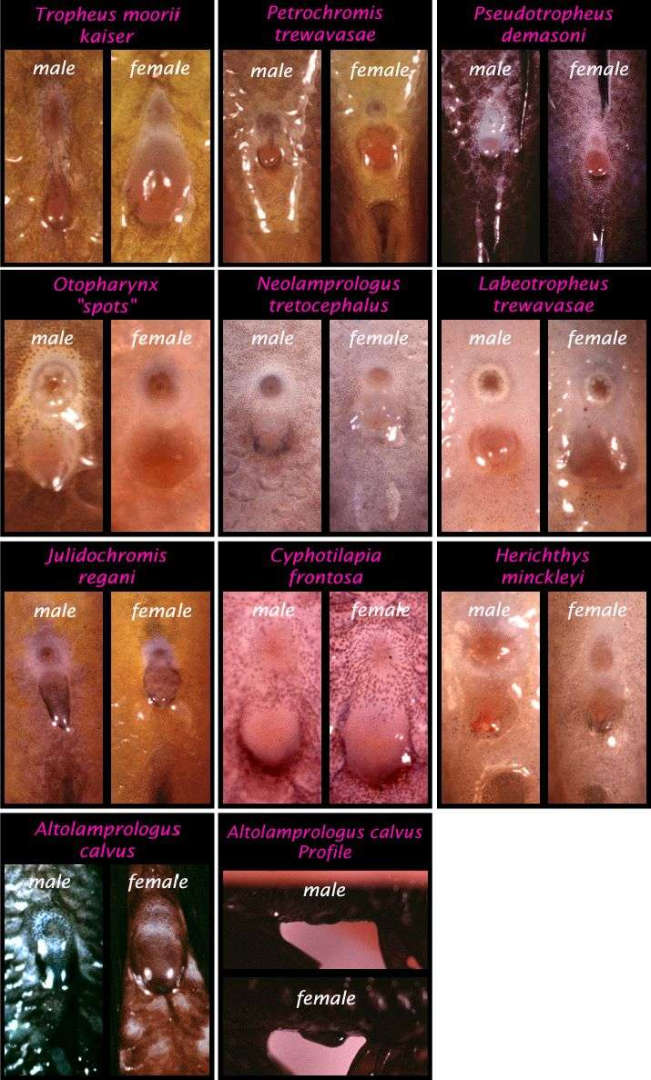
Определение пола у цихлид является одним из самых популярных вопросов. Это не всегда легко или надежно, как хотелось бы, но не смотря на сложность, существует несколько способов определения половой принадлежности рыбок. Если аквариумные рыбки проявляют диморфизм, то есть самец и самка имеют различную окраску, задача упрощается. Все, что нужно это найти фотографии особей данного вида и сравнить с ними. Иногда диморфные рыбки обладают мельчайшими отличиями, что затрудняет определение пола. В данном случае решение загадки с половой принадлежностью особи ведется также как и для мономорфного вида, у которого самцы и самки выглядят одинаково. Определение пола особей мономорфного вида сводится к рассмотрению их наружных половых органов («venting»). Эта процедура заключается в отлове рыбы, помещения её дорсальной стороной книзу и изучения генитального отверстия между анальным отверстием и анальным плавником. Рострально (спереди) расположенное отверстие это анус, а следом за ним идет генитальная папилла (овипозитор у самок). Генитальная папилла, либо овипозитор представляют собой пору, через которую рыбки выделяют икринки или молоки. В общем случае, самка имеет больший овипозитор по сравнению с генитальной папиллой самца. Это связано с приспособлением к выделению более крупной икры. Данные различия сложно определить для видов, которые откладывают икринки на субстрат (в частности, представители рода Neolamprologus), потому что их икринки мельче по сравнению с видами, инкубирующими кладку во рту.
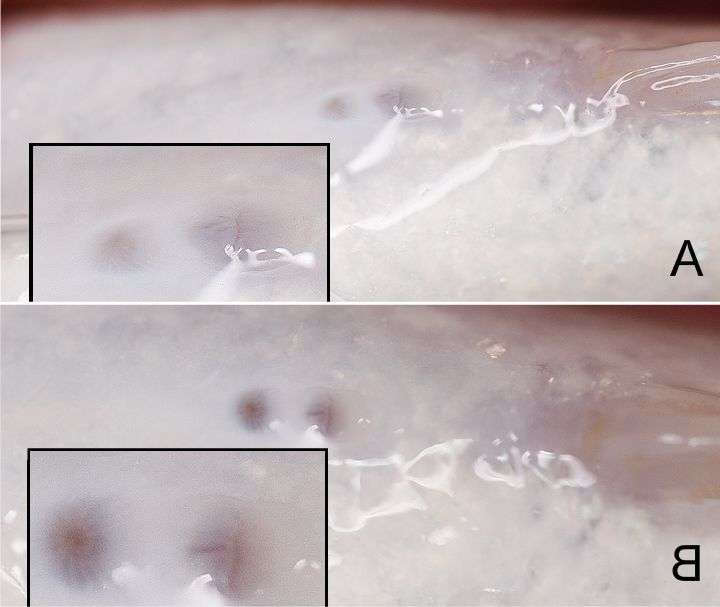
Генитальная папилла танганьиканской цихлиды Limnochromis auritus. У самца (B) анальное отверстие и папилла имеют одинаковый размер,у самки (A) папилла чуть больше ануса (Фото: www.cichlidae.com/forum/viewtopic.php?f=7&t=8720).
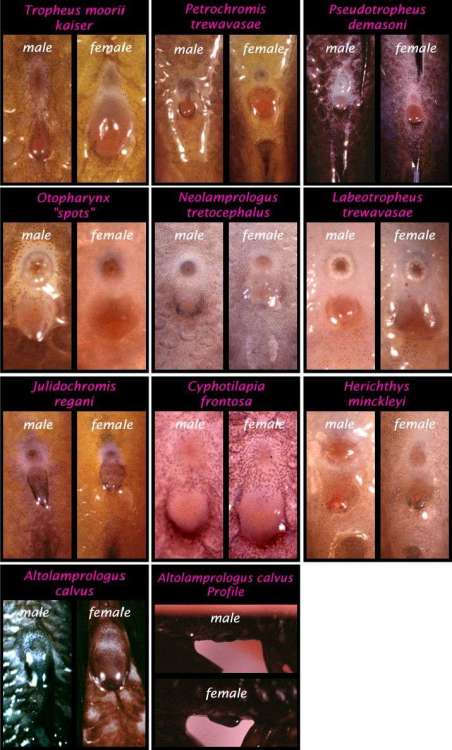

На иллюстрации сравниваются папиллы самцов и самок некоторых видов цихлид.
Сверху располагается анальное отверстие, снизу генитальная папилла или овипозитор (фото: www.fishhead.com/articles/ventsex.htm).
В некоторых случаях представители одного вида демонстрируют различия пигментации полового отверстия у самцов и самок. У некоторых видов, самцы имеют удлиненную и сужающуюся генитальную папиллу, например, у Julidochromis regani. У данных самцов можно провести пальцем через половую пору слева направо, и папилла также будет двигаться. Когда проводится процедура определения пола, необходимо брать рыбку только влажными руками. Фиксация особей через сачок или сетку добавляет надежности и не дает рыбкам соскользнуть. Нельзя передерживать особей в воздухе более 30 секунд. Весь процесс должен быть как можно менее стрессирующим, рыбок желательно вынимать из воды не более трех раз. Как только пол определен, особь помещается отдельно от других с неопределенной половой принадлежностью.
Существует дополнительное количество косвенных признаков, по которым можно судить о поле цихлиды. К ним относятся различия в поведении самца и самки, различия в размере и форме плавников, а также присутствие у самца релизера на анальном плавнике ("egg spot"). Большинство самцов видов цихлид, вынашивающих кладку во рту, имеют пятна на анальном плавнике, похожие на икринки. К сожалению, видов, у которых лишь самец обладает релизером, крайне мало. Многие самки также имеют, по крайней мере, одно икринковое пятно. Данное обстоятельство делает определение по наличию или отсутствию пятна ненадежным способом, но он также имеет право на существование, потому что самцы, в большинстве случаев, имеют больше икринковых пятен. Немаловажным показателем половой принадлежности является более крупный размер тела самца по сравнению с самкой. В некоторых случаях, в частности, у цихлид озера Малави, форма плавников является относительно надежным способом различения особей. В заключении хочется отметить, что для африканских цихлид наилучший метод определения пола сводится к рассмотрению наружных половых органов. Особи, достигшие половой зрелости, будут легче распознаваться, но с опытом аквариумист может научиться с высокой точностью отличать пол большинства видов при возрасте особей менее 4 месяцев.ССЫЛКИ
http://aquavitro.org/2011/09/26/opredelenie-pola-cixlid/
http://www.tanganyika.ru/articles/article.php?art_id=127&cat=1&igen=1
-
41 минуту назад, Stizos сказал:
Свет-хороший свет "спасет отца русской демократии"
-
Melanochromis mpoto, Konings et al, 2012,

Название вида происходит от слова mpoto которое на языке Чи-Тумбуку (наречие на котором разговаривают племена на северо-востоке озера), означает "северный", что характеризует распространение вида в озере Малави.
Первое описание вида сделано в работе : Konings, Ad &, Jay R. Stauffer Jr. 2012. "Review of the Lake Malawi genus Melanochromis (Teleostei: Cichlidae) with a description of a new species". Zootaxa. v. 3258, pp. 1-27
Melanochromis 'blue', Ribbink et al, 1983, временное название
Melanochromis robustus, Konings, 1995, ошибка идентификации
Melanochromis benetos, Schraml, 1998, ошибка идентификации
Melanochromis spec. ‘Matema’, Schraml, 1998, временное название
Melanochromis mpoto, Konings et al, 2012, оригинальное название
Взрослые самцы (В): Голова и тело голубые. Спинной плавник голубой с бледно-голубым краем и белыми концами. Хвостовой плавник голубой с бледно-голубой субмаржинальной полосой и узким желтым краем. Анальный плавник серого или синего цвета с голубым краем и 3-5 желтыми пятнами релизерами. Брюшной плавник серый или синий с белым или голубым передним краем. Грудной плавник со светло-серыми лучами и прозрачными мембранами. В полной окраске самцы окрашены в синий цвет и темные полосы едва различимы, у субдоминантных особей, подростков и самок вертикальные темные полосы выделены довольно ярко.
Самки (С): голова коричневая или темно-коричневая с серой или коричневой окраской основного фона жаберной области. Тело голубовато-коричневое / темно-коричневое, часто с более темными горизонтальными и вертикальными полосами. Спинной и хвостовой плавники серо-коричневые с широким желтым задним краем; анальный плавник серый или коричневый с 1-3 мелкими пятнами релизерами. Брюшной плавник коричневый с узким белым или светло-голубым передним краем. Грудной плавник с коричневыми лучами и серо-желтыми мембранами.
Ареал
Типовая локация: Остров Катале (Katale Island) Малваи, Озеро Малави, , 10° 27.3' S, 34 17.2'.
Melanochromis mpoto имеет довольно широкое распространение в водах озера, в северо-западной части озера от острова Читанде (Chitande Island), около Чилумба (Chilumba) и залива Нката (Nkhata Bay), и в северо-восточной части озера от Матема (Matema) и острова Хонги (Hongi Island) до Лиули (Liuli). Есть информация Риббинка который встречал популяции вида в районе острова Ликома (Likoma Island), но дальнейшие исследования это не подтверждают. -
-
-
Haplochromis sp. 'Cochon Isl.'
Эти фотографии представляют собой совершенно новый вид из озера Киву, абсолютно неизвестный науке, и пойманный ранее в 2018 году совместными усилиями Оливера Лукануса и Ассоциации Цихлид Новой Англии (NECA). Пара на фотографии была поймана у небольшого острова в северном районе озера Киву - Остров де Кохон, и именно так мы называем этих прекрасных хаплохромин до тех пор, пока они не будут официально описаны. Мы получили 6 диких рыб, а Пол Визер, сделавший первую фотографию, сохранил их и размножил. Мы распространили их среди 4 других членов NECA в США, и мы продолжаем находить хорошие разводни для наших подростков и мальков. Эти фотографии дикой рыбы, сделал Оливер Луканус около 6 месяцев назад, и вчера была сделана единственная фотография самца. В целом прекрасное дополнение к нашему хобби.Haplochromis sp. 'Cochon Isl.'
Peter George


-
-
Астаксантин гавайский масляный(лучше усваивается)
Ингредиенты: Природный астаксантин (из микроводорослей гематококкуса плювиалис). Витамин Е (d-альфа-токоферол) жирорастворимый
Также содержит: естественно встречающиеся каротиноиды, включая лютеин, в синергетической смеси.
Без: дрожжей, кукурузы и пшеницы. Без ГМО, без BSE и без глютена и лактозы.

-
1 час назад, Алекс73 сказал:
пошто животину тиранишь?

-
-
-
31 минуту назад, Ефимян сказал:
Как говорил один товарищ - Много фото не бывает!Камаконды ,паразиты,хороши
 .Встречу в Новосибе ,возьму,пока вот не встречал.А вот с юликами не понял,у тебя фотик врёт по цвету ,или они действительно такого цвета?
.Встречу в Новосибе ,возьму,пока вот не встречал.А вот с юликами не понял,у тебя фотик врёт по цвету ,или они действительно такого цвета?
реально такого там подсветка немного в красный LED пришлось даже красный убирать в фш
-
3 часа назад, Ефимян сказал:
Да тут пожалуй этому фото и место.Но Серёг,я человек капризный,хотелось бы всё таки чего нибудь лично от Сергея Аникштейна,так сказать из залежей.
уже давно не кальвусы уже были и марксмита и камаконде бюшери всех развел и даже помногу))) ну покажу что нибудь..много..ничего??











-
Evert van Ammelrooy
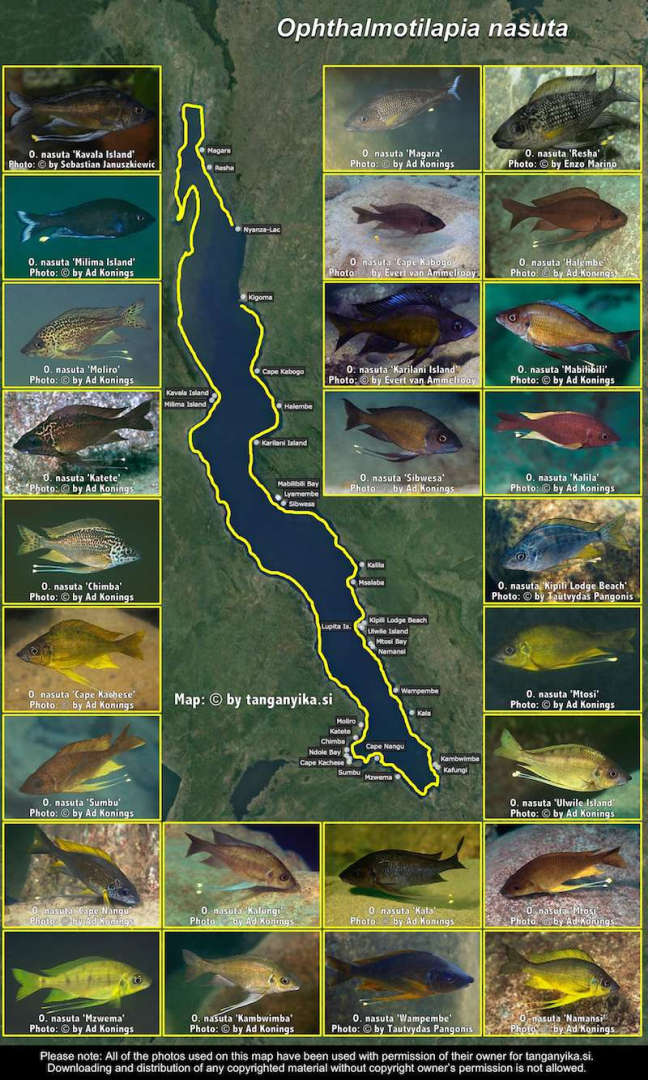
Ophthalmotilapia nasuta 'Kipili Lodge Beach'.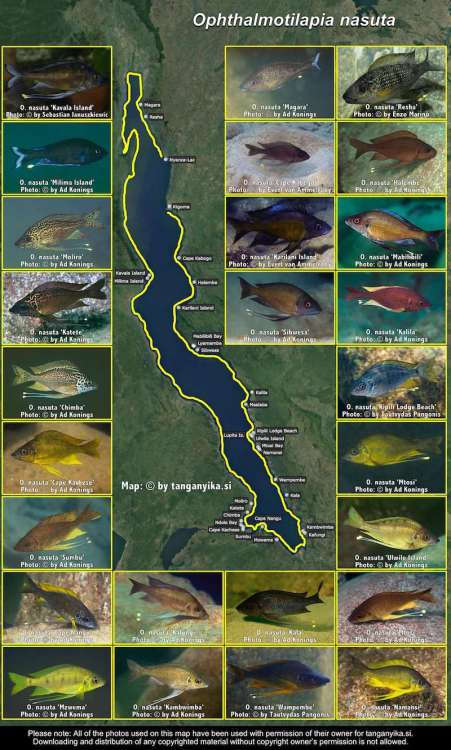


-
Изменение пола у Metriaclima cf. livingstoni в результате влияния поведенческих факторов.
Jay R. Stauffer, Jr., Renea A. Ruffing

Смена пола в естественных условиях, часто встречающаяся у морских рыб, редкое явление у рыб пресноводных. В аквакультуре весьма распространено явление смены пола у Тиляпий путем воздействия на них стероидными препаратами. Было замечено, что сексуальная ориентация этих цихлид достаточно изменчива по сравнению с другими видами и может легко меняться под воздействием стероидных гормонов (Nakamura, 1981). В дикой природе, среди пресноводных рыб, ранее зарегистрирован факт смены пола у Monopterus albus (угря, обитающего в рисовых полях) (Chan, Phillips, 1967). Среди морских же обитателей такие случаи достаточно часты, например, у представителей подотряда губановидных (Labroidei), при этом по мнению многих ученых цихлиды очень схожи с ними по своему строению и происхождению. О смене пола именно у малавийских цихлид упоминается на примере Melanochromis auratus (Hale и др., 1999). Однако для того, чтобы научно доказать, что цихлиды озера Малави способны регулировать соотношение мужских и женских особей путем смены пола, необходимо либо детальное изучение рыб, находящихся в «переходном» состоянии, либо прямое визуальное наблюдение за продуктивным нерестом одной и той же особи до и после смены пола. Целью этого исследования является определение экспериментальным путем возможности смены пола у Metriaclima cf. livingstoni.
ЧИТАТЬ ПОЛНОСТЬЮ http://www.malawi.cichlids.ru/articles/article.php?&art_id=356&cat=12&igen=1
-
В естественных условиях смена пола это довольно частое явление среди морских рыб, у которых преобладает трансформация самцов в самок . В данном случае пол определяется рядом генетических факторов, а также факторами среды, например, pH и температурой. После наступления половозрелости поведенческие взаимодействия могут запустить начало смены пола, то есть наблюдается последовательный гермафродитизм. В большинстве случаев особи, получившие во время взросления один пол, затем необратимо меняют его на противоположный. У протогенных видов особи трансформируются от самок в самцов, у протандрических — первостепенным полом является мужской, который затем сменяется некоторыми рыбками на женский. Одна из гипотез, объясняющая целесообразность гермафродитизма, в качестве ключевого аспекта рассматривает размер тела. Согласно ей, рыбки меняют пол в зависимости от ожидаемой прибавки в размерах. Как правило, плодовитость самцов, в частности, количество продуцируемых гамет, не зависит от их длины и массы. То есть самцы любой длины имеют одинаковую плодовитость. Икринки значительно крупнее сперматозоидов, отсюда, чем больше самка, тем она более плодовита. Будучи маленьким и молодым, самец в полной мере проявляет свой репродуктивный потенциал, но достигнув критического размера превращается в самку (протандрический вид). У протогенных рыбок трансформация пола призвана максимизировать размеры самцов, что объясняется важностью охраны гарема и своей территории. У некоторых гонохористичных (гонохоризм — раздельнополость) рыбок, у которых пол четко дифференцирован на протяжении всей жизни, половые отличия начинают проявляться уже на ранней стадии развития. Однако среди гонохор имеются представители, являющиеся самками на раннем этапе онтогенеза, а затем при посредничестве генетических факторов, трансформирующиеся в самцов . Часто после изменения пола в гонадах у самцов данных видов остаются ооциты.
Изначальная представленность популяции рудиментарных гермафродитов только самками навела на мысль, что женский пол является полом "по-умолчанию". Даже протандрические рыбки изначально не дифференцируются как самцы. В частности, гонады неполовозрелого Клоуна меланопуса (Amphiprion melanopus — Pomacentridae) содержат преимущественно женскую ткань. Тестикулярные элементы начинают преобладать во время созревания особей в самцов, однако они исчезают при трасформации в самку. Подобный механизм был обнаружен и у других видов рода Amphiprion и у аквариумных экземпляров Sparus aurata (Sparidae). Кроме того, у молоди протандрических видов семейства Creediidae обнаружены межполовые гонады, позднее созревающие преимущественно в мужские. Таким образом, мужской пол протадрических видов является скорее переходной, нежели начальной стадией трансформации. Хотя гермафродитизм обычен для рифовых морских рыб, он крайне редко встречается у пресноводных видов. В природе естественная смена пола последними плохо изучена, поэтому причисление некоторых из них к животным с дихогамией (последовательный гермафродитизм) является скорее исключением из правил. Одним из первых, документально зафиксированным случаем смены пола в естественной среде обитания можно назвать транформацию особей вида Monopterus albus, рисового угря . Среди пресноводных обитателей самыми исследуемыми в вопросе половой дифференцировки являются Цихлиды. Это связано с чрезвычайным разнообразием стратегий размножения, включающих откладывание икры на субстрат, либо инкубацию во рту опеку за потомством одним , обоими родителями , групповую или даже межвидовуюзаботу за молодью, сложную систему брачных связей (моногамия, полигамия, стратегия ухода).
Множество работ описывают различные проявления половой лабильности данного семейства пресноводных рыбок. Одним из первых является исследование Полдера, который в 1971 году на примере Aequidens portalegrensis открыл феномен смены пола у южно-американских цихлид . Это породило гипотезу о возможности смены пола у других цихловых. Она была подкреплена Питерсом (1975) ], который при проведении гистологических исследований обнаружил в гонадах самцов 14 видов малавийских цихлид как ооциты, так и сперматогонии. Данные экспериментов свидетельствуют о том, что у изученных представителей группы Мбуны наблюдается гермафродитизм, чаще проявляющийся в трансформации самца в самку. Позднее, Запанс (1985) заявил, что особи вида Cynotilapia afra, члена группы Мбуна, также изменяют пол от самца к самке. Однако данное исследование не включает специальной проверки заявляения. Полевые наблюдения Рибинка и коллег (1983)продемонстрировали, что все члены 200 близких видов группы Мбуна озера Малави вынашивают икру во рту. Но среди некоторых представителей Pseudotropheus lombardoi (Барджесс, 1977) вынашивают икру особи, имеющие окраску самцов. Имеется неколько возможных объяснений этого феномена, включающие вынашивание икры взрослым самцом, самкой, имеющей как самцовую, так и обычную окраску, либо смену пола без сопутствующих изменений окраски. При проведении поведенческих исследований с последующим анатомическим вскрытием ]. было показано, что в семенниках всех самцов находятся недоразвитые ооциты. Это говорит о прохождении гонад Pseudotropheus lombardoi через интерсекуальный ювенильный период. На поздней стадии развития рыбок, тестикулярные элементы доминируют в гонадах, хотя последние и содержат незрелую ткань ооцитов. Самки Pseudotropheus lombardoi проявляют диморфизм, когда некоторые из них имеют сине-желтую окраску, а другие традиционную синюю. Кроме анатомических перестроек у трансформированных самок наблюдаются поведенческие реакции, аналогичные самцам, например, охрана территории и строительство гнезда. Половая система трансформированных самок не притерпевает изменений, в гонадах присутствуют ооциты на различных стадиях развития, отсутствует атрезия ооцитов и развитие ткани семенников.
Таким образом, самки не утрачивают детородную функцию, но при прочих равных, они пользуются гораздо меньшим предпочтением со стороны самцов по сравнению с обычными самками. Более сложная картина транформации наблюдалась при изучении малавийского вида Metriaclima cf. livingstoni . В эксперименте самца отделяли от трех самок с помощью прозрачного пластика. После удаления самца, у одной из самок развивались самцовые вторичные половые признаки, и она начинала оплодотворять икру двух других особей. Во втором эксперименте 6-7 функциональных самок помещалось в аквариум, и в отсутствии самца вновь происходила смена пола самки, которая приступала к нересту и оплодотворяла икру. Аналогичные результаты получены для вида Crenicara punctulata, который по результатам поведенческих исследований и изучению гистологии половой системы трансформированных особей признан протогенным. [4]. Таким образом, при рассмотрении разнообразия цихлид можно отметить присутствие у некоторых видов предрасположенности к смене пола (фенотипической или полной).
Малая распространенность последовательного гермафродитизма у данного семейства и возможно других пресноводных рыб, вероятно, объясняется невозможностью прогнозирования присутствия пищи и полового партнера по сравнению с коралловыми рыбами. Так, у морских рыб очень распространено поддержание постоянных мест кормления, которые особи охраняют. По наблюдениям Гранта (1997) постоянные кормовые угодья наблюдаются у 67% морских коралловых рыб и лишь у 6% морских рыб вод восточной Канады и 9% пресноводных рыб Канады. Благодаря изобилию особей-потенциальных половых партнеров в местах кормления, пища формирует социальную структуру . Тем не менее, различия в защите кормовых угодий не настолько сильные: 84% коралловых рифовых рыбок охраняют эти области, а также места нереста и свое потомство; тоже самое делают 73% морских рыб восточной Канады и 68% пресноводных . Кроме того, спаривание на коралловых рифах обычно происходят продолжительное время, тогда как тропические пресноводные рыбки нерестятся сезонно . Короткие периоды размножения, обычные для озерных и речных рыб, а также отсутствие постоянных областей кормления не позволяют предсказывать распределение половых партнеров. Однако гипотеза, включающая размерную модель (см. выше), нуждается в точной оценке будущего репродуктивного потенциала. То есть основой для формирования последовательного гермофродитизма служит определенность (места и времени) или прогнозируемость предстоящего спаривания.
-
извини тезк забыл где твоя тема с интересными моментами..скину сюда пока
Serkan Odabaşıoğlu

-
8 часов назад, rombao сказал:
Васи особо этим грешат, у меня также отофаринкса 5-6см заглотил гаденыш
васи хищники..это нормально а вот аписто постом выше-это нонсенс

















Neolamprologus wauthioni (Poll, 1949) существует !
в Цихлиды оз. Танганьика
Опубликовано
Neolamprologus wauthioni (Poll, 1949)
Исходное описание Lamprologus wauthioni:
Poll, Max. 1949. "Deuxième série de Cichlidae nouveaux recueillis par la mission hydrobiologique belge en Lac Tanganyika (1946-1947)". Bulletin de l'Institut Royal des Sciences Naturelles de Belgique.
Номенклатурная история:
• Lamprologus wauthioni, Poll, 1949, оригинальное название
• Neolamprologus wauthioni, Colombé et al., 1985, новая комбинация
Обитаемые страны:
• Конго, Демократическая Республика (эндемик).
Этимология: Посвящается г-ну R. Wauthion, мессионеру в Бельгийском Конго.
Описание: Neolamprologus wauthioni - карликовый лампрологус, похожий по размеру и форме на обитающих в раковинах представителей группы видов Neolamprologus ocellatus, с которыми его долго путали. Однако, как утверждает Бюшер (2007), он однозначно отличается несколькими признаками, среди которых его меньшее количество анальных шипов (6 вместо 7-9), удлиненные анальные плавники (против округленных у оцеллатуса) и большее количество позвонков (31 против 29).
Как доказал Бюшер, ближайшим родственником Neolamprologus wauthioni, вероятно, является Neolamprologus ventralis, обитающий у того же побережья, хотя и более южнее. Оба вида отличаются меньшим количеством чешуек в средней линии N. wauthioni (30-31 против 34-36), разным количеством жаберных лепестков (6-7 против 4-5) и разной формой хвостового плавника (округлые у N. wauthioni).
Размер: максимальный известный размер составляет 53 мм.
Половой диморфизм: неизвестен, самцы, возможно больше, чем самки.
Тип населенного пункта: между Albertville и Katibili, в 1 км от берега озера Танганьика.
Распространение. Сохранившиеся экземпляры, являющиеся единственными известными экземплярами, поступают с западного конголезского побережья к югу от Albertville (ныне Калемие) в двух областях: в 15 км к югу и в 35 км к югу (Рутуку).
Место обитания: вид был пойман на довольно большой глубине (35 м), вероятно, над промежуточной или песчаной зоной с улитками Neothauma и их раковинами.
Питание в природе. В кишечнике более крупного обследуемого экземпляра(53 мм) содержались мелкие ракообразные и песок. Как и большинство других лампрологинов, Neolamprologus wauthioni, вероятно, в основном питаются беспозвоночными.
Размножение: нет данных, безусловно, размножается на субстрате, возможно копает норы. Но поскольку типы были собраны с улитками Неотаума, этот вид мог быть случайным обитателем раковин.
Аквариумистика: Никогда не содержался, предположительно похож в содержании на своего вероятного ближайшего родственника, Neolamprologus ventralis.
Сохранение: Neolamprologus wauthioni оценивается международным союзом по сохранению природы в Красном списке угрожаемых видов из за отсутствия данных (DD) (2006).
Комментарии: Идентификация Neolamprologus wauthioni, которую долго путали с членом группы видов Neolamprologus ocellatus (главным образом Neolamprologus speciosus), является хорошим примером того, как небольшая путаница может привести к ошибкам десятилетий. Как подробно доказал Бюшер (Büscher, 2007), этот вид был предварительно оценен по сравнению с членами группы '.L' ocellatus (в основном из-за сравнений Poll), а затем позже с уверенностью рассматривался как один из них опытными специалистами по цихлидам, исключая наиболее очевидное доказательство, а именно то, что члены группы '.L' ocellatus имеют очень своеобразные округлые анальные плавники, в то время как у Neolamprologus wauthioni они одни из самых удлиненных из всех лампрологинов. Эта путаница настолько укоренилась в сознании каждого, что некоторые биомолекулярные исследования были ошибочно основаны на типах «.L» ocellatus, а должны были представлять Neolamprologus wauthioni. Хотя теперь кажется, что N. wauthioni, вероятно, является родственником Neolamprologus ventralis(Büscher), обитающим в той же прибрежной зоне и который, очевидно, никогда не импортировался.